表皮生长因子受体(EGFR)配体
表皮生长因子受体(egfr)配体
1.相关申请的交叉引用
2.本技术要求于2019年7月10日提交的新加坡专利申请第10201906403t号的优先权,出于所有目的将其全部内容通过引用并入本文。
技术领域
3.本发明属于肽/蛋白质技术领域,具体涉及一种稳定性提高的egf受体配体及其方法和用途。
背景技术:
4.慢性伤口和严重疼痛、截肢和残疾的伴随后果仍然是一个重要的健康问题,并增加了社会经济负担。至少有15%的糖尿病患者会在脚上出现慢性溃疡。在美国,治疗这种疾病的总医疗费用估计为90-130亿美元。在这方面,在增殖、迁移和分化中起重要作用的生长因子有望作为伤口管理的治疗干预。表皮生长因子(egf)是1960年发现的第一个生长因子,已在多个国家获得临床批准用于治疗糖尿病足溃疡。然而,egf在慢性伤口的微环境中的蛋白水解稳定性较差,需要频繁施用才能达到预期的治疗效果。最近的研究结果表明,与单独使用egf相比,联合蛋白酶抑制剂局部施用egf可以改善伤口愈合。
5.表皮生长因子受体(egfr/erbb1/her)是一种主要的酪氨酸激酶受体(rtk),可启动发育生长所需的多种细胞反应。表皮生长因子(egf)是第一个发现的egfr激动剂。从那时起,在几乎所有形式的生物体中都发现了egfr激动剂,例如哺乳动物、病毒、昆虫和线虫,但在植物中却没有。它们包括七种相关的哺乳动物egfr激动剂,涵盖53到132个氨基酸残基,其中三个是高亲和力激动剂:egf、转化生长因子-α(tgf-α)和肝素结合性表皮生长因子(hb-egf)。其余四种,β动物纤维素、双调蛋白、上皮调节蛋白和epigen,是低亲和力激动剂。它们都具有一致的egf样结构域。迄今为止,egfr激动剂都有典型的egf结构域,无一例外。由于这些激动剂与rtk的erb家族不同成员的复杂相互作用,它们会产生不同的生物学反应。egfr配体也可以在非哺乳动物来源中找到,例如痘病毒(疫苗生长因子、粘液瘤病毒生长因子和shope纤维瘤病毒生长因子)、秀丽隐杆线虫(lin-3)和果蝇(spitz)。
6.egfr配体通过依赖配体的egfr激活而在细胞增殖、存活和分化中发挥重要作用。所有已知的egfr肽激动剂都具有进化上保守的egf样结构域,由半胱氨酸基序、二硫键连接(cys i-iii、cys ii-iv、cys v-vi)和具有双链反向平行β-折叠的三环结构组成。egf被引入临床环境中用于再生医学,但其应用受到其体内稳定性低的限制。
7.迄今为止,egf是许多治疗糖尿病足溃疡的药物和用于帮助皮肤年轻化的化妆品中的主要活性成分,2016年估计在护肤行业的市场规模为1.02亿美元,平均年增长率为6.27%。由于其作为蛋白质的性质,egf的缺点是它非常容易受到各因素的影响(例如温度和与其他试剂,特别是蛋白酶的相互作用)而影响其稳定性。因此,需要一种egf替代物,其与egf相比表现出(蛋白水解和温度)稳定性提高同时保持与egfr的高亲和力结合和活化egfr。
8.植物是生物活性化合物的丰富来源,对药物设计和开发具有重要价值。然而,由于人民普遍认为其具有不稳定性,因此植物肽和蛋白质往往未被充分探索。
9.本发明通过提供可用于各种应用的超稳定、来源于植物的egfr配体来满足这一需求,这些应用包括皮肤和伤口应用(包括伤口愈合、皮肤和组织再生)以及用于细胞扩增和提高细胞存活率的细胞农业/食品应用(包括培养肉)。
技术实现要素:
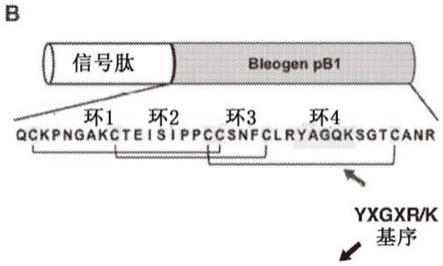
10.本发明基于发明人鉴定出源自植物pereskia bleo的新型富含半胱氨酸的肽(crp),这些肽具有egfr结合活性和高稳定性。富含半胱氨酸的肽(crp)通常是小肽,其特征是半胱氨酸含量高,并且通过二硫桥交联。它们的构象受限结构和富含半胱氨酸的核心为它们提供了相对较高的稳定性。据发现药用植物pereskia bleo是富含半胱氨酸的肽的丰富来源,这些肽统称为bleogens。原型和阳离子bleogen pb1是具有36个残基的肝素结合性crp,其中5个是lys/arg,6个是cys,半胱氨酸基序为c(x)6c(x)7cc(x)3c(x)
10
c(seq id no:2)。与egfr配体在结构上的不同之处在于,它具有与典型的egf样结构域不同且更加紧凑的结构。bleogen pb1采用四环结构,其二硫键以胱氨酸结连接方式排列(cys i-iv、cys iiv和cys iii-vi)。bleogen pb1生物合成为双结构域前体,成熟结构域在信号肽酶裂解后释放。bleogen pb1具有有助于其肝素结合特性的阳离子-极性-阳离子基序。bleogen pb1还与扭结素型(knottin-type)抗微生物肽具有序列同源性并表现出抗念珠菌特性。因此,bleogen pb1代表了在发现egf的59年后的一流的egfr激动剂。
11.在第一方面,本发明因此涉及具有egfr结合活性的分离肽,所述肽包含或由以下组成:
12.(i)seq id no:1中列出的氨基酸序列;
13.(ii)与seq id no:1中列出的氨基酸序列在其整个长度上具有至少60%,优选至少70%,更优选至少80%,最优选至少90%序列同一性的氨基酸序列;
14.(iii)与seq id no:1中列出的氨基酸序列在其整个长度上具有至少80%,优选至少90%,更优选至少95%序列同源性的氨基酸序列;或者
15.(iv)(i)-(iii)中的任一片段。
16.由seq id no:1组成的肽在本文中也称为“bleogen pb1”或“pb1”。
17.另一方面,本发明还涉及编码本文所述的肽的核酸分子,以及含有这种核酸的载体,特别是复制载体或表达载体。
18.在另一方面,本发明还涉及宿主细胞,优选非人宿主细胞,其包含如本文所考虑的核酸或如本文所考虑的载体。宿主细胞可以是细菌细胞,例如大肠杆菌,或植物细胞。
19.本发明的另一方面是一种如本文所述的肽的生产方法,包括培养本文所考虑的宿主细胞;从培养基或宿主细胞中分离肽。另一方面涉及通过化学合成(例如固相肽合成)来生产如本文所述的肽的方法。
20.在另一方面,本发明涉及包含本文所述的肽的组合物,特别是药物、化妆品或药妆组合物。所述组合物还可包含载体和/或赋形剂。
21.在另一方面,本发明涉及本文所述的肽或含有它们的组合物用于激活egfr的用途,特别是在离体应用例如细胞培养中的用途。
22.在另一方面,本发明涉及一种或多种本发明的肽或本发明的组合物用于预防或治疗有需要的受试者的egf或egfr相关疾病或病症的方法。该方面还涵盖本发明的肽或化妆品/药物/药妆组合物在制备用于治疗或预防有需要的受试者的egf或egfr相关疾病或病症的药物中的用途,其中所述预防或治疗可包括施用美容、治疗或预防有效量的本发明的肽或组合物。
23.在另一方面,本发明涉及治疗或预防有需要的受试者的egf或egfr相关疾病或病症的方法,包括向该受试者施用美容、预防或治疗有效量的一种或多种本发明的化合物或本发明的组合物。
附图说明
24.图1.bleogen pb1和七种相关哺乳动物egfr激动剂的序列和结构比较。(a)pereskia bleo的叶子;(b)bleogen pb1的前体结构、一级序列和二硫键连接;(c)weblogo显示的所有七种相关哺乳动物egfr激动剂(egf、tgfα、hb-egf、β细胞素、双调蛋白、上皮调节蛋白和epigen)的序列比较;yxgxr基序(x,任何氨基酸)以红色阴影表示。(d)bleogen pb1的环4与egf的环c(pdb条目:1p9j)、上皮调节蛋白(pdb条目:1k36)和tgfα(pdb条目:1yuf);(e)使用cluspro 2.0版服务器对bleogen pb1和egfr细胞外域(pdb条目:1ivo)之间的相互作用进行建模。
25.图2.bleogen pb1的化学合成。(a)bleogen pb1和生物素pb1的合成方案,在wang树脂上使用逐步固相fmoc化学法。bleogen pb1是在wang树脂上使用逐步固相fmoc化学合成的,以产生pg-pb1(pg:保护基团)。生物素-pb1是通过将fmoc-lys(生物素)偶联到树脂上pg-pb1的n端来合成的。tfa裂解后,从树脂载体释放的组装线性前体立即在ph8.0的0.1m碳酸氢铵和10%二甲亚砜(dmso)中进行氧化折叠1小时,其中氧化还原试剂半胱胺/胱胺的摩尔比为10:1;(b)使用rp-hplc洗脱天然的和合成的bleogen pb1;(c)天然的和合成的bleogen pb1使用肝素亲和色谱法的hplc色谱图。
26.图3.bleogen pb1发挥egf样活性。(a,b)天然的和合成的bleogen pb1依赖剂量促进hacat细胞增殖72小时(通过结晶紫检测),以egf作为阳性对照;所有结果均表示为三个独立实验的平均值
±
标准差;(c)bleogen pb1促进原代人角质细胞增殖72小时(使用结晶紫检测),以egf作为阳性对照;所有结果均表示为三个独立实验的平均值
±
标准差;与对照相比,*p《0.05;(d)bleogen pb1在体外促进ha-cat细胞中的dna合成(通过edu掺入测试),以egf作为阳性对照;所有结果均表示为三个独立实验的平均值
±
标准差;与对照组相比,*p《0.05。(e)连续5天皮下注射bleogen pb1(3mg/kg,10只小鼠)目视检查到新生小鼠的门牙萌出从153小时(盐水对照,10只小鼠)加速到125小时,而阳性对照egf(3mg/kg,10只小鼠)将门牙萌出加速至100小时;门牙萌出被定义为给定牙齿第一次刺穿口腔上皮的时间;n=每组10只小鼠;所有结果均表示为平均值
±
标准差;与对照组相比,*p《0.05。(f)在使用c57小鼠的5mm全厚度夹板切除伤口模型(总共50处伤口;25只小鼠)中,与盐水载体对照组(n=30处伤口;15只小鼠)相比,受伤后连续三天局部应用bleogen pb1(10nmol/伤口;n=10处伤口;5只小鼠)和egf(1nmol/处伤口;n=10处伤口;5只小鼠)加速了伤口愈合。上图:处理方案的示意图。下图:受伤后14天的过程中伤口愈合的百分比。所有结果均表示为平均值
±
标准差;与盐水对照组相比,所有处理组的*p《0.05。
27.图4.bleogen pb1与egfr结合。(a)使用抗egfrmab进行hacat细胞裂解物的egfr的生物素-pb1、生物素-ab1(阴性对照)和生物素-egf(阳性对照)拉下实验的代表性免疫印迹图像;n=三个独立实验;(b)抗egfr中和的mab(克隆la1)阻断了bleogen pb1在hacat细胞中的增殖作用(通过结晶紫检测)。小鼠igg1用作对照;所有结果均表示为三个独立实验的平均值
±
标准差;与pb1处理组相比,*p《0.05。
28.图5.bleogen pb1激活egfr及其下游信号通路。(a)在5μmbleogen pb1培养后免疫沉淀样品使用egfr抗体(偶联磁珠)对hacat细胞中磷酸化酪氨酸(p-tyr)和egfr的代表性免疫印迹分析;n=三个独立实验;(b)在5μmbleogen pb1培养后对磷酸化mek1/2(p-mek1/2)、总mek(t-mek1/2)、磷酸化erk1/2(p-erk1/2)和总erk1/2(t-erk1/2)在hacat细胞中的表达的代表性免疫印迹分析;n=三个独立实验;(c,d)ag1478(一种egfr特异性酪氨酸激酶抑制剂)和u0126(一种mek特异性抑制剂)在无血清培养基中72小时对bleogen pb1诱导的hacat细胞增殖的影响。所有结果均表示为三个独立实验的平均值
±
标准差;与pb1处理组相比,*p《0.05。(e)bleogen pb1培养6小时会增加稳定转染的sre-荧光素酶报告基因hacat细胞中的荧光素酶活性。egf用作阳性对照。所有结果均表示为三个独立实验的平均值
±
标准差;与对照组相比,*,#p《0.05。(f)bleogen pb1处理2小时上调了hacat细胞中c-fos和c-jun的基因表达。所有结果均表示为三个独立实验的平均值
±
标准差;与对照组相比,*,#p《0.05。
29.图6.使用d-氨基酸对bleogen pb1中的yagqk区域进行位置扫描。
30.(a)使用不同浓度的bleogen pb1、d-类似物([y25y]pb1、[a26a]pb1、[q28q]pb1、[k29k]pb1)、ab1(阴性对照)、rt7(阴性对照)或egf(阳性对照)进行基于tr-fret的egfr的生物素-egf竞争性置换。所有结果均表示为三个独立实验的平均值
±
标准差;(b)使用hacat细胞的1μmbleogen pb1或d-类似物([y25y]pb1、[a26a]pb1、[q28q]pb1、[k29k]pb1)增殖效应72小时(通过结晶紫检测)。所有结果均表示为三个独立实验的平均值
±
标准差;与对照组相比,*p《0.05。与pb1组相比,#p《0.05。(c)抗egfr中和的mab(克隆la1)阻断了[k29k]pb1在hacat细胞中的增殖作用(通过结晶紫检测)。小鼠igg1用作对照;所有结果均表示为三个独立实验的平均值
±
标准差;与[k29k]pb1组相比,#p《0.05。与[k29k]pb1和抗egfrmab组相比,*p《0.05。
[0031]
图7.bleogen pb1和[k29k]pb1加速链脲佐菌素诱导的糖尿病小鼠的伤口愈合。在使用stz诱导的糖尿病c57小鼠的5mm全厚度夹板切除伤口模型(总共48处伤口;24只小鼠)中,与盐水载体对照组(n=12处伤口;6只小鼠)相比,在受伤后连续5天局部施用bleogen pb1(1nmol/伤口;n=12处伤口;6只小鼠)、[k29k]pb1(1nmol/伤口;n=12处伤口;6只小鼠)和egf(1nmol/伤口;n=12处伤口;6只小鼠)加速了伤口愈合。上图:处理方案的示意图。下图:受伤后14天的过程中伤口愈合的百分比。所有结果均表示为平均值
±
标准差;与盐水对照组相比,所有处理组的*p《0.05。
[0032]
图8.bleogen pb1和[k29k]pb1是超稳定egfr激动剂。通过rp-hplc分析的bleogen pb1、[k29k]pb1、egf和s-烷基化pb1(碘乙酰胺-)在(a)热(100℃)、(b)人血清、(c)胃蛋白酶、(d)胰蛋白酶、(e)链霉蛋白酶,和(f)中性粒细胞弹性蛋白酶处理下的稳定性;所有结果均表示为三个独立实验的平均值
±
标准差;n.d.:未检测到。
[0033]
图9.使用肠激酶从mbp-pb1融合蛋白中去除mbp的质谱图。
[0034]
图10.通过反相hplc(rp-hplc)共洗脱天然和重组bleogen pb1。
具体实施方式
[0035]
本发明基于发明人鉴定出分离自pereskia bleo的、具有egfr结合活性的新型富含半胱氨酸的肽(crp)。具体而言,发明人成功地从pereskia bleo中鉴定了一种名为bleogen pb1的新型超稳定egfr配体。原型和阳离子bleogen pb1是具有36个残基的肝素结合性crp,其中5个是lys/arg,6个是cys,半胱氨酸基序为c(x)6c(x)7cc(x)3c(x)
10
c(seq id no:2)。与之前已知的egfr配体结构不同,bleogen pb1采用四环结构,其二硫键以胱氨酸结连接方式排列(cys i-iv、cys ii-v和cys iii-vi)。bleogen pb1生物合成为双结构域前体,成熟结构域在信号肽酶裂解后释放。bleogen pb1具有有助于其肝素结合特性的阳离子-极性-阳离子基序。bleogen pb1还与扭结素型(knottin-type)抗微生物肽具有序列同源性并表现出抗念珠菌特性。
[0036]
发明人还发现,bleogen pb1不含egf样结构域,因此是在发现egf的59年后的一流的egfr激动剂。36个残基的bleogen pb1是迄今为止报道的最小的天然肽基egfr激动剂。它比两种最小的egfr激动剂分别短10个和16个残基,即46个残基的上皮调节蛋白和52个残基的tgf-α。egf和tgf-α被归类为高亲和力egfr激动剂,而双调蛋白和上皮调节蛋白被归类为低亲和力激动剂。受体置换试验表明,bleogen pb1的效力比egf低约50-100倍,使其成为低亲和力egfr激动剂。与egf类似,bleogen pb1促进角质形成细胞增殖、角质形成细胞迁移。bleogen pb1的环4与tgf-α的环c具有高度的序列同一性和结构相似性。存在保守的yxgxk/r(seq id no:3)基序表明它们具有共同的egfr相互作用“热点”。
[0037]
以前的研究(us 5,182,261;tam j&ke x(1989)systematic approach to study the structure-activity of transforming growth factorα.peptides:chemistry and biology[proceedings of the 11th american peptide symposium],(escom,leiden),pp 75-77)已经表明,tgf-α的yxgxr环c中的突变,特别是y38和r42,导致其在a431细胞中的egfr亲和力和egf样有丝分裂效力显着降低。同样,在对egf的单独研究中,其y37和r41突变类似物显示其egf样活性降低(ogiso et al.(2002)crystal structure of the complex of human epidermal growth factor and receptor extracellular domains.cell110(6):775-78746,62-65)。总的来说,这些发现表明yxgxr基序作为egfr激动剂的推定的受体接触位点的重要性。
[0038]
通过点置换bleogen pb1中的yagqk基序,可以证明所有ala置换的pb1类似物的egf样生物活性降低,这与egf和tgf-α的观察结果一致。相比之下,两种d-氨基酸置换的pb1类似物k29k pb1和y25y pb1显示出egf样生物活性增加。在k29k pb1中通过用d-lys替换第29位的lys,低亲和力bleogen pb1转化为高亲和力egfr激动剂。据发现k29k pb1与egf一样有效,其中与bleogen pb1相比,效力提高了60倍。体外结果得到了体内伤口愈合stz糖尿病小鼠模型的支持,该模型表明k29k pb1和egf的作用相当。
[0039]
序列比对表明,bleogen pb1的k29对应于egf的r41和tgf-α的r42。该位置的arg是一个关键残基,在所有已知的典型egfr激动剂中绝对保守(ogiso,同上)。结构和突变研究还确定了egf的r41残基对于与egfr的d355形成盐桥是关键的,对于受体结合是必不可少的(ogiso,同上)。与先前对tgf-α和egf的突变研究一致,这些结果表明k29是bleogen pb1的
egf样活性的重要的分子决定因素。
[0040]
与egfr激动剂相比,bleogen pb1具有相似的功能特征,但其一级序列、二级和三级结构以及生物合成不同。所有已知的egf家族激动剂,尤其是哺乳动物来源的激动剂,都包含egf样结构域(singh,carpenter&coffey(2016)egf receptor ligands:recent advances.f1000research 5)。此外,哺乳动物egfr激动剂生物合成作为1型跨膜前体,并在被解离素和金属蛋白酶(adam)的蛋白水解裂解后释放。相比之下,bleogen pb1不包含egf样结构域;相反,它包含seq id no:2的半胱氨酸基序,这是6-半胱氨酸-橡胶蛋白(hevein)样肽家族的典型特征,并且被生物合成为由er信号肽和成熟结构域组成的双结构域前体。这种半胱氨酸基序是一种胱氨酸结二硫键连接,比egf更能抵抗蛋白水解降解。因此,bleogen pb1是一类新的超稳定egfr激动剂的原型成员,具有非典型的一级序列、二级和三级结构、生物合成途径和高蛋白水解稳定性。
[0041]
已经通过开发基于聚合物的系统、封装和纳米技术致力于延长egf的半衰期。据发现bleogen pb1和有效的k29k pb1显示出egf样活性,并共同具有结构紧凑的支架,该支架比egf对蛋白水解降解的稳定性至少高100倍。将lys从其l型替换为d型产生k29k pb1,其效果与egf相当。通过进一步开发当前的方法,扩大修饰肽(如k29k pb1)的生产是可行的。利用正交trna/合成酶配对可以简化非天然氨基酸的掺入,以在大肠杆菌中进行生物合成生产(liu&schultz(2010)adding new chemistries to the genetic code.annu.rev.biochem.79:413-444;liu et al.(2012)genetic incorporation of d-lysine into diketoreductase in escherichia coli cells.amino acids 43(6):2553-2559)。此外,对时间和成本高度有利的化学合成和片段缩合方法可用于工业制造。使用这些技术的市售例子有比伐红素(一种凝血酶抑制剂)和t-20(一种hiv融合抑制剂)。总之,bleogen pb1是一种超稳定的、非典型的、最小的来源于植物的egfr激动剂,它的发现是一个前景很好的开局。改进的pb1类似物k29k pb1与egf同等有效这一发现可以进一步推动用于伤口愈合、再生医学和护肤的有效治疗类似物的开发。
[0042]
基于上述发现,本发明在第一方面涵盖分离形式的具有活性的肽,并且更具体地涉及分离的肽,其包含seq id no:1(bleogen pb1)中列出的氨基酸序列或基本上由seq id no:1(bleogen pb1)中列出的氨基酸序列组成。
[0043]
由seq id no:1中列出的氨基酸序列组成的肽在本文中也称为“bleogen pb1”或“pb1”。如本文所用,“分离”是指肽的形式已与它可能天然存在或与之相关的其他细胞成分至少部分分离。肽可以是重组肽,即在不天然产生所述肽的基因工程生物体中产生的肽。
[0044]
根据本发明的肽表现出egfr结合活性,即它能够以特定方式识别和结合egfr,即优先于其他受体类型而与之结合,通常比非特异性结合所观察到的亲和力高至少10倍或100倍。此外,肽优选还表现出egfr活化活性,即用作egfr激动剂。在各种优选的实施方案中,egfr结合类似于egf的结合,即亲和力在egf的
±
50%以内。
[0045]
如本文所用,“肽”涉及由通过肽键连接的氨基酸得到的聚合物。如本文所定义的肽可包含10个或更多个氨基酸,优选20个或更多个,更优选25个或更多个氨基酸,例如25至50个氨基酸,更优选30至40个或32至36个氨基酸。如本文所用,“多肽”涉及包含超过100个氨基酸的肽。
[0046]
在各种实施方案中,肽包含或由以下组成:与seq id no:1中列出的氨基酸序列在
其整个长度上具有至少60%、65%、70%、75%、76%、77%、78%、79%、80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、90.5%、91%、91.5%、92%、92.5%、93%、93.5%、94%、94.5%、95%、95.5%、96%、96.5%、97%、97.5%、98%、98.5%、99%、99.25%或99.5%序列同一性或同源性的氨基酸序列。或者,同一性或同源性可以与seq id nos.12-14中的任一个有关。在一些实施方案中,它具有与seq id no:1、12、13或14中列出的氨基酸序列在其整个长度上具有至少60%、优选至少70%、更优选至少80%、最优选至少90%序列同一性的氨基酸序列,或者具有与seq id no:1、12、13或14中列出的氨基酸序列在其整个长度上具有至少80%、优选至少90%、更优选至少95%序列同源性的氨基酸序列。
[0047]
在各种实施方案中,肽可以是成熟酶的前体。在此类实施方案中,除了在seq id no:1、12、13或14中列出的那些氨基酸序列之外,它还可包含额外的氨基酸序列。此类前体典型地可包含n端信号肽,其典型长度为20-30个氨基酸,可能在翻译后加工过程中被切割。
[0048]
核酸序列或氨基酸序列的同一性通常通过序列比较来确定。这种序列比较基于现有技术中建立并常用的blast算法(参见例如altschul et al.(1990)“basic local alignment search tool”,j.mol.biol.215:403-410,和altschul et al.(1997):“gapped blast and psi-blast:a new generation of protein database search programs”;nucleic acids res.,25,p.3389-3402),原则上分别使核酸序列和氨基酸序列中相似的核苷酸或氨基酸的连续序列相互关联。相关位置的表格关联被称为“比对”。通常使用本领域技术人员可用和已知的计算机程序来生成序列比较(比对),特别是多序列比较。
[0049]
这种类型的比较还允许报告被比较的序列彼此之间的相似性。这通常表示为百分比同一性,即在比对中在相同位置或彼此对应的位置的相同核苷酸或氨基酸残基的比例。在氨基酸序列的背景下,术语“同源性”更广泛地解释为也包括考虑保守氨基酸交换,即具有相似的化学活性的氨基酸,因为它们通常在蛋白质内表现出相似的化学活性。保守氨基酸置换包括但不限于:本发明上下文中的保守氨基酸置换包括,例如,g=a=s、i=v=l=m、d=e、n=q、k=r、y=f,s=t,a=i=v=l=m,y=f=w。因此,比较序列的相似性也可以表示为“百分比同源性”或“百分比相似性”。同一性和/或同源性的指示可以在整个(多)肽或基因上遇到,或仅在个别区域上遇到。因此,通过序列中的匹配来定义各种核酸序列或氨基酸序列的同源区域和相同区域。这些区域通常表现出相同的功能。它们可以很小,并且只包含几个核苷酸或氨基酸。这种小区域通常执行对蛋白质整体活性至关重要的功能。因此,将序列匹配仅参考个别区域和可选的小区域可能很有用。然而,除非另有说明,本文中同一性和同源性的指示是指分别指示核酸序列或氨基酸序列的全长。
[0050]
在多个实施方案中,本文所述的肽在对应于seq id no:1的2、9、17、18、22和33位的任何一个或多个位置处包含氨基酸残基c,优选至少在2和18、9和22和/或17和33位置,更优选前述对中的两组,最优选所有六处位置。
[0051]
在各种实施方案中,至少存在富含半胱氨酸的核心基序c(x)nc(x)mcc(x)oc(x)
p
c,其中x可以是除c之外的任何氨基酸,n为4至8的整数,m为5至9的整数,o为1至5的整数,且p为8至12的整数,优选c(x)6c(x)7cc(x)3c(x)
10
c(seq id no:2);和/或氨基酸序列基序yxgxk/r(seq id no:3),其中x可以是除c之外的任何氨基酸,优选地它们组合存在。
[0052]
在各种实施方案中,氨基酸基序yxgxk/r(seq id no:3)可以包含在基序c(x)nc(x)mcc(x)oc(x)
p
c的(x)
p
部分中或者seq id no:2的基序的(x)
10
部分中。因此,在各种实施
方案中,本发明的肽包含氨基酸序列c(x)6c(x)7cc(x)3cxxyxgxk/rxxxc(seq id no:4),ck/rpxgxk/rcxxxxxppccxxxcxk/ryxgxk/rxgxcxxk/r(seq id no:5),或ckpxgxkcxexxxppccxxxcxryxgxkxgxcxxr(seq id no:6)。
[0053]
seq id no:3的基序可以是yagxk(seq id no:15),其中x可以是除c之外的任何氨基酸,优选d、e、n或q,更优选n或q,最优选q。seq id no:3的基序也可以是yxgqk(seq id no:16),其中x可以是除c之外的任何氨基酸,优选a、v、l、s,更优选a、v或s,最优选a。这些实施方案还包括肽,其中seq id no:15或16的基序包括在如上定义的富含c的基序中或seq id no:2、4、5或6中。
[0054]
在各种实施方案中,seq id nos.2-6和15-16中列出的氨基酸保留在seq id no:1的主链中,而剩余的氨基酸可以在给定的序列同一性和/或同源性范围内变化。在这些实施方案中,所述置换可以包括任何其他氨基酸置换为c。
[0055]
在各种实施方案中,肽包含对应于seq id no:1的残基2-33的氨基酸。在各种实施方案中,本发明的片段包含对应于seq id no:1的氨基酸2-33的氨基酸序列。因此,片段的长度优选为至少32个氨基酸。本发明包括的其他片段是对应于seq id no:1或相应序列的n-和/或c-端截短形式的片段。n-端截短的片段优选仅缺少第一个n-端氨基酸(使用seq id no:1的位置编号),而c-端截短的片段可以从c-末端开始缺少1-3个氨基酸(使用seq id no:1的位置编号)。在各种实施方案中,存在对应于seq id no:1的36位的氨基酸可能是有利的。
[0056]
在各种实施方案中,本发明的肽与seq id no:1中列出的序列的不同之处在于它们包含一个或多个氨基酸变异,选自插入、缺失和/或置换。在各种实施方案中,seq id no:1的这些变体包含seq id nos.2-6和15-16中的任何一个或多个。在各种实施方案中,本发明的肽包含一个或多个氨基酸置换。这些置换可以包括一个天然存在的l-氨基酸被另一个l-氨基酸置换,优选如本文公开的选择,特别是在seq id nos.2-6和15-16中的任一个中指定为x的位置。在其他实施方案中,置换是l-氨基酸被d-氨基酸置换,通常是相应的d-氨基酸。因此,肽可以包含一个或多个d-氨基酸。在各种实施方案中,这些置换包含y25y和/或k29k,最优选的是包含k29k置换的变体。此类变体的氨基酸序列在seq id nos.7-9中列出。如下定义,单字母代码中大写字母表示l-氨基酸,而单字母代码中小写字母表示d-氨基酸。因此,“k”是l-赖氨酸,而“k”是d-赖氨酸。因此“k29k”意指在对应于seq id no:1中29位的位置处的l-赖氨酸(k)被d-赖氨酸(k)置换。如果没有另外指明,本文所用的所有位置编号是指使用seq id no:1作为参考的位置编号。
[0057]
在各种实施方案中,肽可具有正净电荷。这意味着肽包含氨基酸残基,使得带正电荷的氨基酸(h、k、r)的所有正电荷总和高于带负电荷的氨基酸(d、e)的所有负电荷的总和。换言之,肽中存在的h、k和r的氨基酸残基总和高于氨基酸残基d和e的总和。
[0058]
在各种实施方案中,肽包含一个、两个或三个二硫桥,优选三个,优选地选自c2-c18之间、c9-c22之间以及c17-c33之间的二硫桥,使用seq id no:1的位置编号。
[0059]
在多个实施方案中,本发明的肽对热和/或蛋白酶的稳定性与野生型egf相比(其氨基酸序列在seq id no:10中列出)高至少2倍,优选至少5倍,更优选至少10倍。这些可以使用本领域技术人员已知的常规检测来确定,其中在相同条件下测定egf和本发明的肽。
[0060]
在本文中通常通过单字母代码指代所有氨基酸残基,并且在一些情况下,使用它
们的三字母代码。该命名法是本领域技术人员众所周知的,并且如本领域所理解的那样在本文中使用。置换是指起始氨基酸、位置编号和目标氨基酸。例如,“k29a”是指29位的k被置换为a。此外,本文中以大写字母指代的或没有任何进一步说明的所有氨基酸都是l-氨基酸。在本文中使用单字母代码以小写字母表示d-氨基酸。因此,“k29k”表示29位的l-赖氨酸被d-赖氨酸置换。此外,在本文给出的任何序列中,由“/”分隔的氨基酸意味着那些可以替代地存在。因此,“k/r”是指在给定位置可能存在k或r残基。
[0061]
除了上述修饰之外,根据本文描述的实施方案的肽可以包含氨基酸修饰,特别是氨基酸置换、插入或缺失。例如,通过靶向遗传修饰进一步开发此类肽,即通过诱变方法,并针对特定目的或特殊性质(例如,其活性、稳定性等)进行优化。如果将此类额外修饰引入本发明的肽中,则它们优选不影响、改变或逆转上文详述的序列基序,即富含c的基序(除可变残基x之外)和yxgxk/r环。这意味着这些残基/基序的以上定义的固定位置不会被以上定义之外的这些额外突变改变。
[0062]
在各种实施方案中,本发明的肽可以被翻译后修饰,例如糖基化。此类修饰可通过重组方式进行,即在产生时直接在宿主细胞中进行,或可在多肽合成后通过化学或酶促方式实现,例如在体外。
[0063]
本文所描述的修饰的目的可以是将靶向突变(例如置换、插入或缺失)引入已知分子中,以便例如增加结合特异性/亲和力和/或提高活性。为此,尤其可以改变分子的表面电荷和/或等电点,从而改变它们与靶的相互作用。或者或另外,肽的稳定性可以通过一个或多个相应的突变来增强。个体突变(例如个体置换)的有利特性可以相互补充。此类修饰的实例已在上文中描述,包括将所选氨基酸置换为其d-氨基酸对应物,特别是k29k变体。
[0064]
在各种实施方案中,肽的特征在于其可通过上述多肽作为单个或多个保守氨基酸置换的初始分子而获得。术语“保守氨基酸置换”是指一个氨基酸残基交换(置换)为另一个氨基酸残基,其中这种交换不会导致被交换的氨基酸的位置处的极性或电荷发生变化,例如将一个非极性氨基酸残基替换为另一个非极性氨基酸残基。本发明上下文中的保守氨基酸置换包括例如上文公开的置换。
[0065]
或者或另外,肽的特征在于其可通过本文的肽作为初始分子经过片段化或缺失、插入或置换诱变而获得,并且包含与seq id nos.1、12-14中列出的初始分子在其至少32个连续连接的氨基酸的整个长度上匹配的氨基酸序列(对应于seq id no:1的氨基酸2-33)。优选地,在此类实施方案中,氨基酸c2、c9、c17、c18、c22和c33以及y25、g27和k29仍以其天然l-构象或人工引入的d-形式存在。
[0066]
在各种实施方案中,本发明因此还涉及本文描述的肽的片段,所述片段保留所需的结合和活性。优选地,它们具有初始分子(优选具有seq id no:1的氨基酸序列的肽)的至少50%、更优选至少70%、最优选至少90%的亲和力和/或活性。优选的片段已在上文定义。
[0067]
编码本文所述的肽的核酸分子,以及含有这种核酸的载体,特别是复制载体或表达载体也构成本发明的一部分。
[0068]
这些可以是dna分子或rna分子。它们可以作为单链存在、作为与所述单链互补的单链存在或作为双链存在。特别是对于dna分子,在每种情况下都要考虑所有三个可能的阅读框中的两条互补链的序列。还要考虑的是,不同的密码子(即碱基三联体)可以编码相同的氨基酸,因此特定的氨基酸序列可以由多种不同的核酸编码。由于遗传密码的这种简并
性,可以编码上述肽之一的所有核酸序列都包括在本发明的这个主题中。技术人员能够明确地确定这些核酸序列,因为尽管遗传密码具有简并性,但确定的氨基酸与单个密码子相关联。因此,技术人员可以从氨基酸序列出发,容易地确定编码该氨基酸序列的核酸。此外,在根据本发明的核酸的背景下,一个或多个密码子可以被同义密码子替换。该方面特别是指本文考虑的肽的异源表达。例如,每个有机体(例如生产菌株的宿主细胞)具有特定的密码子用法。“密码子用法”被理解为由相应生物体将遗传密码翻译成氨基酸。如果位于核酸上的密码子在生物体中遇到相对少量的负载trna分子,则蛋白质生物合成中可能会出现瓶颈。它也编码相同的氨基酸,因此密码子在生物体中的翻译效率低于编码相同氨基酸的同义密码子。由于同义密码子存在大量trna分子,因此后者可以在生物体中更有效地翻译。
[0069]
通过目前公知的方法,例如化学合成或聚合酶链反应(pcr)结合分子生物学或蛋白质化学的标准方法,技术人员有能力在已知dna序列和/或氨基酸序列的基础上制造相应的核酸乃至完整的基因。此类方法是已知的,例如从sambrook,j.,fritsch,e.f.,and maniatis,t,2001,molecular cloning:a lab manual,3rd edition,cold spring laboratory press中了解。
[0070]
出于本文的目的,“载体”被理解为由核酸组成的成分,其包含本文考虑的核酸作为特征核酸区域。它们使所述核酸能够在物种或细胞系中经过多代或细胞分裂建立为稳定的遗传成分。特别是当用于细菌时,载体是特殊的质粒,即环状遗传成分。在本文的背景下,如本文所考虑的核酸被克隆到载体中。例如,载体包括起源于细菌质粒、病毒或噬菌体的载体,或者主要是具有广泛不同衍生成分的合成的载体或质粒。使用在每种情况下都存在的其他遗传成分,载体能够在多代相关宿主细胞中将自身建立为稳定单元。它们可以作为单独的单位存在于染色体外,或者可以分别整合到染色体中进入染色体dna。
[0071]
表达载体包括能够在宿主细胞中复制的核酸序列,优选包含它们的微生物,特别优选细菌,并在其中表达所包含的核酸。在各种实施方案中,本文所述的载体因此还包含控制编码本发明多肽的核酸表达的调控成分。表达特别受调控转录的一个或多个启动子的影响。原则上可以通过最初位于待表达的核酸前面的天然启动子发生表达,也可以通过在表达载体上提供的宿主细胞启动子,或者通过修饰的、或完全不同的、另一个生物体或另一个宿主细胞的启动子发生表达。在本发明中,至少一种用于表达本文所考虑的核酸的启动子是可获取的并可用于其表达。此外,表达载体可以被调控,例如通过改变培养条件或含有它们的宿主细胞达到特定细胞密度,或通过添加特定物质,特别是基因表达激活子。这种物质的一个例子是半乳糖衍生物异丙基-β-d-硫代半乳糖吡喃糖苷(iptg),它用作细菌乳糖操纵子(lac operon)的激活子。与表达载体相反,所包含的核酸不在克隆载体中表达。
[0072]
在另一方面,本发明还涉及宿主细胞,优选非人宿主细胞,其包含如本文所考虑的核酸或如本文所考虑的载体。优选地,将本文所考虑的核酸或含有所述核酸的载体转化到微生物中,则其代表根据一实施方案的宿主细胞。在现有技术中已经建立了用于转化细胞的方法并且是技术人员充分已知的。原则上所有细胞都适合作为宿主细胞,即原核或真核细胞。优选可以以遗传有利(例如关于使用核酸或载体的转化,及其构建的稳定性)的方式操作的宿主细胞。此外,优选的宿主细胞以在微生物学和生物技术方面易于操作而著称。例如,这指的是易于培养、高生长速率、对发酵培养基的需求低以及外源蛋白质的良好生产和分泌速率。肽可以在它们制造后通过产生它们的细胞进行修饰,例如通过添加糖分子、甲酰
化、胺化等。这种类型的翻译后修饰可以在功能上影响肽。
[0073]
进一步实施方案的宿主细胞其活性可以基于可获取的遗传调控成分进行调控,例如在载体上、但也可以先天地存在于那些细胞中。例如,可以通过控制添加用作激活子的化合物、改变培养条件或达到特定的细胞密度来刺激它们表达。因此能够经济地生产本文考虑的蛋白质。如前所述,这种化合物的一个例子是iptg。
[0074]
优选的宿主细胞是原核或细菌细胞,例如大肠杆菌细胞。细菌的显着特点是世代时间短,对培养条件的要求很少。因此,可以建立经济的培养方法和制造方法。此外,技术人员在发酵技术中的细菌方面具有丰富的经验。在个别情况下,根据实验确定的多种因素,例如营养来源、产物形成率、时间要求等,革兰氏阴性或革兰氏阳性细菌可能适用于特定的生产实例。在各种实施方案中,宿主细胞可以是大肠杆菌细胞。
[0075]
本文考虑的宿主细胞可以根据它们对培养条件的要求进行修饰,可以包含其他或额外的选择标记,或者也可以表达其他或额外的蛋白质/肽。它们尤其可以是转基因表达多种肽的宿主细胞。
[0076]
然而,宿主细胞也可以是真核细胞,其特征在于它具有细胞核。因此,另一个实施方案为具有细胞核的宿主细胞。与原核细胞相比,真核细胞能够对形成的蛋白质/肽进行翻译后修饰。实例是真菌如放线菌,或酵母如saccharomyces或kluyveromyces或昆虫细胞,如sf9细胞。这可能是特别有利的,例如当与它们的合成相关的蛋白质进行由此类系统可能实现的特定修饰时。真核系统特别与蛋白质合成结合进行的修饰包括例如结合低分子量化合物,例如膜锚或寡糖。在各种实施方案中,宿主细胞是真核细胞。
[0077]
本文考虑的宿主细胞以常规方式培养和发酵,例如在不连续或连续系统中。在前一种情况下,将宿主细胞接种到合适的营养培养基中,经过一段时间的实验确定后,从培养基中收获产物。连续发酵以实现流动平衡而著称,其中在相对较长的时间内,细胞部分死亡但也部分更新,并且形成的肽可以同时从培养基中去除。
[0078]
本文考虑的宿主细胞优选用于生产本文描述的肽。
[0079]
因此,本发明的另一方面是如本文所述的肽的制造/生产方法,包括在允许所述肽表达的条件下培养本文考虑的宿主细胞;从培养基或宿主细胞中分离肽。培养条件和培养基可以由本领域技术人员根据所使用的宿主生物,通过采用本领域已知的常识和技术来选择。例如,肽的表达可以通过使用融合蛋白进行,其中本发明的肽与促进表达/分离/纯化的另一肽/蛋白融合,例如通过亲和层析进行。通常通过用位点特异性蛋白酶处理来加工这种融合构建体,其中该蛋白酶切割表达/亲和标签并因此释放目标肽。
[0080]
另一方面,本发明涉及本文公开的肽用作药物的用途。因此考虑将本发明的化合物用作药物。
[0081]
在另一方面,本发明涉及一种或多种本发明的肽用于预防或治疗有需要的受试者的egf或egfr相关疾病或病症的方法。该方面还涵盖本发明的肽在制造用于治疗或预防有需要的受试者的egf或egfr相关疾病或病症的药物中的用途,其中所述预防或治疗可包括施用治疗或预防有效量的本发明的肽。“egf或egfr相关的疾病或病症”不仅包括由受试者中egf/egfr的异常活性直接引起的疾病和病症,还包括egf和egfr在其中起信号通路作用并受益于egf/egfr活化的疾病和病症。此类病症包括但不限于伤口愈合、溃疡和神经变性疾病,包括但不限于糖尿病性溃疡、胃溃疡、食道溃疡、十二指肠溃疡、烧伤伤口、手术伤口、
压力伤口、化疗引起的伤口、角膜伤口、阿尔茨海默病,以及皮肤疾病。皮肤相关应用包括但不限于皱纹改善、皮肤水合作用、色素沉着预防、皮肤弹性改善、皮肤干细胞分化等。因此,上述治疗包括促进受试者的伤口愈合或治疗任何提及的病症,例如溃疡和神经变性疾病。
[0082]
在另一方面,本发明涉及治疗或预防有需要的受试者的egf或egfr相关疾病或病症的方法,包括向该受试者施用预防或治疗有效量的一种或多种本发明的肽。
[0083]
通常,本发明的肽将通过本领域已知的任何常用和可接受的方式以治疗/预防有效量单独或与一种或多种其他治疗剂联合给药。治疗/预防有效量可根据疾病的严重程度、受试者的年龄和相对健康状况、所用肽的效力和其他因素而广泛变化。
[0084]
本发明的肽可以作为药物组合物通过任何常规途径给药,特别是局部给药,例如以洗剂、凝胶、滴眼液、软膏或霜剂的形式,也可以肠胃外给药,例如以可注射溶液或悬浮液的形式。这样的应用还包括剂型,例如创可贴,其中组合物可以提供在载体材料上,例如织物或纤维材料,以及水凝胶。
[0085]
因此,本发明还涉及包含一种或多种本发明的肽和药学上可接受的赋形剂或载体的药物组合物。载体可包括稀释剂和/或溶剂。
[0086]
可以通过混合、制粒或包覆方法以常规方式制造包含游离形式或药学上可接受的盐形式的本发明的肽以及至少一种药学上可接受的载体或稀释剂的药物组合物。组合物可以被灭菌和/或含有辅助剂,例如防腐剂、稳定剂、润湿剂或乳化剂、溶液促进剂、用于调节渗透压的盐和/或缓冲剂。此外,它们还可能含有其他具有治疗价值的物质。用于例如皮肤和眼睛的局部应用的合适制剂优选是本领域公知的水溶液、软膏、乳膏或凝胶。这些可能含有增溶剂、稳定剂、张力增强剂、缓冲剂和防腐剂。
[0087]
本发明的肽可以以治疗有效量与一种或多种治疗剂联合施用(药物组合)。可与本发明化合物联合使用的化合物的非限制性实例有:已知的蛋白酶抑制剂、egf和egf激动剂。
[0088]
当本发明的肽与其他疗法联合给药时,共同给药的化合物的剂量当然会根据所使用的共同药物的类型、所使用的具体药物、所治疗的病症等而变化。
[0089]
如本文所用,术语“共同给药”或“联合给药”等意在包括将所选治疗剂给药至单个患者,并且旨在包括治疗方案,其中所述药剂的给药途径不一定相同或不一定同时给药。
[0090]
如本文所用,术语“药物组合”是指由一种以上活性成分的混合或组合得到的产品,并且包括活性成分的固定组合和非固定组合。术语“固定组合”是指活性成分(例如本发明的肽和助剂)均以单一实体或剂量的形式同时施用于患者。术语“非固定组合”是指活性成分(例如本发明的肽和助剂)均作为单独实体同步、同时或顺序地施用于患者,没有特定的时间限制,其中这样的施用在患者体内提供了治疗有效水平的两种化合物。后者也适用于鸡尾酒疗法,例如施用3种以上的活性成分。
[0091]
药物组合物可用于预防或治疗有需要的受试者的egf或egfr相关疾病或病症的方法。
[0092]
在另一方面,本发明涉及治疗或预防有需要的受试者的egf或egfr相关疾病或病症的方法,包括向该受试者施用预防或治疗有效量的本发明的药物组合物。
[0093]
上文针对药物组合物描述的所有实施方案类似地适用于也构成本发明的一部分的化妆品或药妆组合物。应当理解,除了本发明的肽之外,存在于这些组合物中的所有伴随剂都是化妆品或药妆可接受的。由于化妆品/药妆组合物通常可以是局部给药形式,因此以
tof ms进行鉴定。使用流动相a(0.1%tfa/h2o)和流动相b(0.1%tfa/乙腈(acn))的线性梯度。通过hplc分析计算折叠产率为70%。进行1h nmr、rp-hplc和肝素亲和色谱以证明合成bleogen pb1与其天然形式相比的完整性。
[0107]
在肽树脂上进行pb1、[y25y]pb1和[k29k]pb1的n端生物素标记,用fmoc-溶素(生物素)-oh(4.0eq.)、n,n-二异丙基乙胺(dipea;6.0eq.)、(苯并三唑-1-基氧基)三吡咯烷鏻六氟磷酸盐(pybop;4.0eq.)在50%二甲基甲酰胺(dmf)和50%1-甲基-2-吡咯烷酮(nmp)中的混合物在室温下进行2小时。反应2小时后,生物素标记的pb1在前面描述的条件下裂解和氧化折叠。折叠的生物素-pb1通过制备型hplc纯化并使用maldi-tof ms进行鉴定。
[0108]
细胞培养和转染
[0109]
hacat(人角质形成细胞)、huvec-cs(内皮细胞)细胞在补充有10%胎牛血清和100u/ml青霉素和链霉素的dulbecco改良的eagle培养基(dmem)中培养。从cell technology公司购买的正常人表皮角质形成细胞在dermalife k培养基中培养。它们在37℃的5%co2加湿培养箱中生长。hacat细胞用pgl4.33[luc2p/sre/hygro](promega)稳定转染,并使用250μg/ml潮霉素进行筛选。用250μg/ml潮霉素维持表达sre荧光素酶报告基因的稳定hacat细胞。
[0110]
细胞增殖试验
[0111]
使用结晶紫染色确定细胞增殖。简而言之,按每孔1.0x 104个将hacat细胞接种在96孔板中,并在无血清培养基中加入bleogen pb1或egf(阳性对照)或3417-da肽ab1(阴性对照)。3417-da-肽ab1是从牛膝分离的crp,氨基酸序列为ncesgtscipgaqhnccsgvcvpivtifygvcy(seq id no:11)。如前所述进行结晶紫染色。简而言之,培养期后,将孔用4%缓冲多聚甲醛固定20分钟。然后将细胞用0.25%结晶紫的20%甲醇染色15分钟。多余的结晶紫染色用蒸馏水漂洗4-5次,风干。加入在milli-q水中的冰醋酸(10%)以提取结晶紫染色。然后使用酶标仪(tecan200pro,瑞士)在595nm处测量吸光度。
[0112]
edu掺入分析
[0113]
根据制造商的说明,使用edu增殖试剂盒(ifluor 488)进行edu掺入试验。简而言之,按每孔2.5x 104个将hacat细胞接种在96孔板中。过夜培养后,细胞与edu(10μm)一起在无血清培养基中与bleogen pb1或egf一起培养4小时。然后将细胞固定、透化并在含有ifluor 488的反应混合物中培养30分钟。使用dapi对细胞核进行染色。使用荧光显微镜观察板。
[0114]
细胞迁移试验
[0115]
使用划痕试验监测细胞迁移。简而言之,将hacat细胞接种到2孔硅胶插件中,间隙为500μm(ibidi,德国)。培养过夜后,取出插件并在含有0.1%fbs的dmem培养基中与bleogen pb1或egf一起培养。培养8小时后,使用倒置相差显微镜对孔进行拍照。使用wimasis图像分析平台分析图像的无细胞区域百分比。
[0116]
内皮管形成试验
[0117]
使用μ-载玻片(ibidi,德国)进行内皮管形成试验。简而言之,将10μl的凝胶基质(bd matrigel
tm basement membrane matrix)浇注到每个μ-载玻片孔上并静置固化。huvec-cs细胞与bleogen pb1或egf一起以2500个细胞/孔的密度接种每个孔。培养2h后,使用倒置相差显微镜对孔进行拍照。使用wimasis图像分析平台分析图像的分支点编号。
[0118]
拉下试验
[0119]
使用neutravidin ultralink树脂(thermo fisher scientific,美国)进行拉下试验。简而言之,将树脂用磷酸盐缓冲盐水(pbs)洗涤3次,并与生物素-pb1/生物素-[k29k]pb1/生物素-[y25y]pb1、或生物素(阴性对照)或生物素-ab1(阴性对照)或生物素-egf(阳性对照)在室温下培养并旋转2小时。将在pbs中的2%牛血清白蛋白(bsa)添加到两个管中,在室温下温育并轻轻地颠倒混合2小时。将600μg hacat细胞裂解液加入每个管中,并在4℃下旋转培养过夜。培养后,将树脂转移至离心柱并用pbs洗涤10次。将含有β-巯基乙醇的6x装载染料加入树脂中,并在85℃下加热10分钟。将所得混合物以200g离心1分钟,然后使用8%十二烷基硫酸钠聚丙烯酰胺凝胶电泳(sds-page)在100v恒定电压下分离120分钟。印迹转移在聚偏二氟乙烯(pvdf)膜(ge healthcare,瑞典)上以250ma在冰上进行120分钟。印迹用5%bsa tris缓冲盐水和吐温20(tbst)封闭,然后在4℃下与抗egfr抗兔抗体(1:1000在5%bsa tbst中;cell signaling,美国)培养过夜。培养过夜后,在室温下用tbst洗涤膜3次,每次10分钟。然后将印迹与二级抗兔辣根过氧化物酶(hrp)(cell signaling,美国)(1:5000在5%bsa tbst中)一起培养,并在室温下再培养1小时。将印迹在室温下用tbst洗涤5次,每次10分钟,然后再添加化学发光底物(advansta,美国)和在x射线胶片(fujifilm,日本)上曝光。
[0120]
免疫沉淀
[0121]
使用与磁珠偶联的egfr(d38b1)兔单克隆抗体(cell signaling,美国)进行免疫沉淀。简而言之,将磁珠用pbs洗涤3次,然后将50μg hacat细胞裂解物加入每个管中,并在4℃下旋转培养过夜。培养后,磁珠用pbs洗涤5次。将含有β-巯基乙醇的6x装载染料加入树脂中,并在85℃下加热10分钟。将所得混合物以14000g离心1分钟,并使用10%sds-page在100v恒定电压下分离120分钟。印迹转移在pvdf膜(ge healthcare,瑞典)上以250ma在冰上进行120分钟。印迹用5%bsa tbst封闭,然后在4℃下与抗磷酸酪氨酸-hrp(r&d systems,美国)或抗egfr抗兔抗体(1:1000在5%bsa tbst中;cell signaling,美国)一起培养过夜。培养后,在室温下用tbst洗涤膜3次,每次10分钟。然后将印迹与二级抗兔辣根过氧化物酶(hrp)(1:5000在5%bsa tbst中)一起培养,并在室温下再培养1小时。将印迹在室温下用tbst洗涤5次,每次10分钟,然后再添加化学发光底物(advansta,美国)和在x射线胶片(fujifilm,日本)上曝光。
[0122]
免疫印迹分析
[0123]
在冰上以250ma 120分钟将印迹转移到pvdf膜(ge healthcare,瑞典)。印迹用5%bsa tbst封闭,然后在4℃下与抗p-mek1/2兔抗体(1:2000在5%bsa tbst中;cell signaling,美国)、抗mek1/2兔抗体(1:2000在5%bsa tbst中;cell signaling,美国),抗p-erk1/2兔抗体(1:2000在5%bsa tbst中;cell signaling,美国),抗erk1/2兔抗体(1:2000在5%bsa tbst中;cell signaling,美国)和抗β-肌动蛋白小鼠抗体(1:10000在5%bsa tbst中;merck millipore,美国)。培养后,在室温下用tbst洗涤膜3次,每次10分钟。然后将印迹与二级抗小鼠或抗兔辣根过氧化物酶(hrp)(1:5000在5%bsa tbst中;cell signaling,美国)一起培养,并在室温下再培养1小时。将印迹在室温下用tbst洗涤五次,每次10分钟,然后再添加化学发光底物(advansta,美国)和在x射线胶片(fujifilm,日本)上曝光。
[0124]
tr-fretegfr配体结合试验
[0125]
根据制造商的说明,使用egf-egfr lance ultra tr-fret结合试剂盒(perkinelmer,美国)进行竞争性置换试验。链霉亲和素与lance铕螯合物结合,其中后者与生物素-egf结合,而egfr-fc与用ulight
tm
染料标记的抗人igg相互作用。简而言之,将不同浓度的bleogen pb1或其ala-或d-氨基酸类似物或egf与工作溶液混合,并在室温下培养2小时。tr-fret比率使用酶标仪在双发射模式下测量(激发:340nm,发射:665nm和615nm)(cytation 1,美国)。结果表示为生物素-egf的相对结合百分比。氨基酸序列为ncesgtscipgaqhnccsgvcvpivtifygvcy(seq id no:11)的ab1或氨基酸序列为cvssgivdacseccepdkciimlptwppryvcsv(seq id no:17)的roseltide rt7用作阴性对照。egf(seq id no:10)用作阳性对照。
[0126]
荧光素酶报告基因试验
[0127]
测试了表达sre荧光素酶报告基因的稳定hacat细胞对用bleogen pb1和egf作为阳性对照处理的生物发光反应。简而言之,将细胞在白色96孔板中培养以达到90%的汇合度。细胞用无血清培养基洗涤,在无血清dmem培养基中饥饿过夜,并用bleogen pb1和egf处理6小时。根据制造商的说明,使用one-glo ex萤光素酶测定系统(promega,美国)进行萤光素酶测定。使用酶标仪(tecan200pro,瑞士)测量发光。
[0128]
基因表达分析
[0129]
使用purelink
tm rna mini试剂盒(thermo fisher scientific,美国)从hacat和原代人角质形成细胞中提取总rna。根据制造商的说明,使用superscript
tm ii逆转录酶和oligo(dt)12-18(thermo fisher scientific,美国)从600ng总rna合成第一链cdna。使用itaq universal sybr green supermix(bio-rad,美国)在cfx96 touch实时pcr检测系统(bio-rad,美国)上进行40个循环的定量pcr(qpcr)。pcr反应(20μl):3μl cdna、1μl引物混合物(10μm)、6μl depc处理水和10μl反应混合物(master mix)。qpcr反应中使用的预先设计的引物对(origene,美国)如下:c-fos(nm_005252)是5'-gcc tct ctt act acc act cac c-3'(正向;seq id no:18)和5'-aga tgg cag tga ccg tgg gaa t-3'(反向;seq id no:19);c-jun(nm_002228)是5'-cct tga aag ctc aga act cgg ag-3'(正向;seq id no:20)和5'-tgc tgc gtt agc atg agt tgg c-3'(反向;seq id no:21);参考基因gapdh(nm_001256799)是5'-gtc tcc tct gac ttc aac agc g-3'(正向;seq id no:22)和5'-acc acc ctg ttg ctg tag cca a-3'(反向;seq id no:23)。gapdh用作标准化的看家基因。使用2-δδct
方法计算c-fos和c-jun基因表达的倍数变化。
[0130]
小鼠全层夹板切除伤口
[0131]
c57bl/6小鼠获自vital rital laboratories(中国北京)。将小鼠关在23
±
1℃的塑料笼中,12小时光/暗循环,提供自由饮水和食物。所有实验均经过批准并按照中国医学科学院实验动物中心(中国北京)的机构指南进行。
[0132]
使小鼠形成全层5毫米夹板切除伤口。简而言之,使用戊巴比妥钠(50mg kg-1
)腹腔注射麻醉小鼠。后背的毛发先用电动推剪剃掉,然后再涂上脱毛膏。使用5毫米直径的无菌活检穿孔器,形成对称的全层切除伤口。使用胶水在伤口周围放置夹板,并用四根间断缝合线固定。伤口和夹板用tegaderm(3m,美国)覆盖。在受伤后14天的指定天数,测量伤口直径。以沿x和y轴的两个直径测量值的平均值计算伤口的大小。
[0133]
c57bl/6小鼠全层夹板切除伤口愈合的处理方案
[0134]
如上所述,对c57bl/6小鼠进行全层5毫米夹板切除伤。受伤后,连续三天用盐水(载体对照;n=30处伤口;15只小鼠)、egf(1nmol/伤口,阳性对照;n=10处伤口;5只小鼠)和bleogen pb1(10nmol/伤口;n=10处伤口;5只小鼠)对伤口进行处理。
[0135]
链脲佐菌素诱导的糖尿病c57bl/6小鼠全层夹板切除伤口愈合的处理方案
[0136]
在第1天和第5天通过两次腹膜内注射stz(80mg/kg)诱导c57bl/6小鼠患上糖尿病。从第8天开始,定期监测血糖两周。血糖水平高于11.1mmol/l的小鼠被认为患有糖尿病,并如上所述进行全层5毫米夹板切除伤(总共24只小鼠,每只小鼠有两处伤口)。受伤后,连续五天将生理盐水(载体对照;n=12处伤口)、bleogen pb1(1nmol/伤口;n=12处伤口)、[k29k]pb1(1nmol/伤口;n=12处伤口)和egf(1nmol/伤口;n=12处伤口)局部施用于伤口。还在受伤后第14天监测血糖以确保小鼠患有糖尿病。
[0137]
新生小鼠门牙萌出模型
[0138]
icr小鼠获自vital rital laboratories(中国北京)。将小鼠关在23
±
1℃的塑料笼中,12小时光/暗循环,提供自由饮水和食物。所有实验均经过批准并按照中国医学科学院实验动物中心(中国北京)的机构指南进行。从出生之日(第0天)开始,在新生小鼠的颈背连续五天进行皮下注射。将总共30只新生小鼠随机分为如下三个测试组:pbs、egf(3mg/kg)和bleogen pb1(3mg/kg)。每天通过目测记录门牙萌出。门牙萌出被定义为给定牙齿首次刺穿口腔上皮的时间。
[0139]
肽稳定性试验
[0140]
热稳定性
[0141]
在100℃下培养0.1m纯化的bleogen pb1、[k29k]pb1、s-烷基化pb1(碘乙酰胺-)、egf、pbs(对照)。在不同的时间点(0、30、60和120分钟)收集样品。
[0142]
酸稳定性
[0143]
将0.1m纯化的bleogen pb1、[k29k]pb1、s-烷基化pb1(碘乙酰胺-)和egf溶解在0.2m hcl中,并在37℃下培养。在不同的时间点收集样品。
[0144]
胃蛋白酶稳定性
[0145]
将0.1m纯化的bleogen pb1、[k29k]pb1、s-烷基化pb1(碘乙酰胺-)和egf溶解在0.2m hcl中,并与胃蛋白酶(roche applied science,美国)以50:1(w/v)的比例在37℃下培养。在不同的时间点收集样品。
[0146]
链霉蛋白酶稳定性
[0147]
将0.1m纯化的bleogen pb1、[k29k]pb1、s-烷基化pb1(碘乙酰胺-)和egf溶解在pbs中,并与链霉蛋白酶(0.2mg/ml;roche applied science,美国)在37℃下培养。在不同的时间点收集样品。
[0148]
中性粒细胞弹性蛋白酶稳定性
[0149]
将0.1m纯化的bleogen pb1、[k29k]pb1、s-烷基化pb1(碘乙酰胺-)和egf溶解在pbs中,并与人中性粒细胞弹性蛋白酶(0.05mg/ml;molecular innovations,美国)在37℃下培养。在不同的时间点收集样品。
[0150]
胰蛋白酶稳定性
[0151]
将0.1m纯化的bleogen pb1、[k29k]pb1、s-烷基化pb1(碘乙酰胺-)和egf溶解在
pbs中,并与胰蛋白酶(0.2mg/ml;sigma aldrich,美国)在37℃下培养。在不同的时间点收集样品。
[0152]
人血清稳定性
[0153]
在不含酚红的dmem培养基中的25%人血清中制备0.1m纯化的bleogen pb1、[k29k]pb1、s-烷基化pb1(碘乙酰胺-)和egf。测试样品在37℃下培养。在不同的时间点收集样品。收集的样品用100%乙醇进行蛋白质沉淀,并在4℃下以180000g离心5分钟。收集上清液用于分析。
[0154]
稳定性试验分析
[0155]
从各种稳定性试验中收集的所有样品均通过rp-hplc进行分析,其中在aeris肽xb-c18柱(phenomenex,美国)上,使用流动相a(0.05%tfa/h2o)和流动相b(0.05%tfa/acn)的线性梯度。通过maldi-tof ms收集和鉴定所得峰。使用hplc曲线的峰面积将结果表示为初始浓度的百分比。
[0156]
统计分析
[0157]
使用graphpad version 8.2.1(us)进行统计比较。使用单向方差分析(anova)和newman-keuls事后检验分析数据。数据表示为平均值
±
标准差。p《0.05被认为具有统计学意义。
[0158]
实施例1:作为egfr激动剂的bleogen pb1的基序搜索和分子对接,以确定构效关系研究的“热点”[0159]
七种相关的哺乳动物egfr激动剂包括egf、tgf-α、hb-egf、β细胞素、双调蛋白、上皮调节蛋白和epigen,共同拥有四个保守的非半胱氨酸残基,其中三个位于环c(cys v-vi)中,形成特定的yxgxr基序(x,任何氨基酸)(图1c)。bleogen pb1的环4也包含一个yxgxk基序,在序列和结构上类似于egfr激动剂的环c(图1d)。使用蛋白质-蛋白质对接服务器cluspro 2.0版对bleogen pb1和egfr(pdb条目1ivo,链a)进行的计算机模拟表明,bleogen pb1环4可以在与egf环c相同的位点与egfr结合,表明这是一个常见的“热点”以结合egfr(图1e)(ogiso,同上)。重要的是,pb1和egfr激动剂共同拥有相同“热点”这一发现为我们随后的构效关系研究提供了依据。
[0160]
实施例2:bleogen pb1的合成和表征
[0161]
对于egf样活性的体外和体内测定,使用从pereskia植物中分离的天然bleogen pb1。从pereskia bleo的叶子提取物中分离天然bleogen pb1,使用c-18反相高效液相色谱(rp-hplc)(loo et al.(2017)bleogens:cactus-derived anti-candida cysteine-rich peptides with three different precursor arrangements.front.plant sci.8:2162)。为了制备合成bleogen pb1,使用了逐步固相法和fmoc化学法(图2a)。在去除保护基团并通过三氟乙酸(tfa)从树脂载体上切割未保护的肽后,使用由半胱胺和胱胺以10:1摩尔比在0.1m碳酸氢铵中组成的氧化还原试剂组合氧化折叠粗pb1产物,ph 8保持1小时以产生70%的bleogen pb1。经rp-hplc、肝素亲和色谱和二维核磁共振(nmr)证明,无法区分纯化的合成和天然bleogen pb1(图2b、c)。
[0162]
为了给bleogen pb1配备用于目标识别的化学亲和探针(图2a),通过将fmoc-lys(生物素)偶联到仍附着在树脂支持物上的受保护的pb1肽的n端,再次全合成,以提供生物素化-pb1(生物素-pb1)。这些全合成为bleogen pb1在其n端提供了明确的位点特异性标
记,以保持其侧链官能团的完整性。
[0163]
此外,还使用iptg诱导表达的lb培养基(0.4mm,37℃下4小时;0.1mm,15℃下20小时;数据未示出)在大肠杆菌中重组产生了bleogen pb1,作为mbp(麦芽糖结合蛋白)融合蛋白。使用麦芽糖树脂通过亲和层析分离表达的融合蛋白,并用10mm在pbs中的麦芽糖洗脱以进行纯化。mbp-pb1的纯化产量约为10-15mg/l lb肉汤。如质谱分析所示,通过用肠激酶处理去除mbp标签(图9)。图10示出天然和重组bleogen pb1使用rp-hplc时在相同的保留时间共洗脱。
[0164]
实施例3:bleogen pb1显示出egf样生物活性
[0165]
为了确定bleogen pb1是否是egf样有丝分裂原,使用egf作为阳性对照检查了其对hacat角质形成细胞增殖的生物学影响。结果表明,bleogen pb1和egf促进hacat细胞增殖,ec50分别为130nm和1.2nm(图3a、b)。egf和pb1之间100倍的促有丝分裂效力差异表明bleogen pb1属于低亲和力egfr激动剂家族,例如epigen和双调蛋白。天然和合成的bleogen pb1对hacat细胞显示出相同的增殖活性,证实了观察到的增殖效应不是由于来自植物提取物的污染物(图3b)。作为阴性对照,从achyranthes bidentata中分离的3417-da肽ab1是一种与bleogen pb1属于相同crp家族的6chlp,具有相似的半胱氨酸基序和二硫键连接,其在该测定中的活性高达10μm。结果还表明bleogen pb1促进原代人角质形成细胞的增殖(图3c),并通过5-乙炔基-2'-脱氧尿苷(edu)掺入试验增强hacat细胞的dna合成(图3d)。此外,egf和bleogen pb1均加速hacat细胞迁移和内皮细胞管形成(数据未示出)。标志性egf试验是新生小鼠的门牙萌出,通过该试验揭露了这一发现。结果表明,连续5天皮下注射bleogen pb1(3mg/kg)或egf(3mg/kg)可加速新生小鼠的门牙萌出,从153小时(盐水对照)分别加速到125小时和100小时(图3e)。结果还表明bleogen pb1显示出体内伤口愈合。在c57小鼠中使用全层切除伤口模型,连续三天使用bleogen pb1(10nmol/伤口)或egf(1nmol/伤口)处理加速了伤口愈合,从第3天到第11天(图3f)(wang et al.(2013),nat.protoc.8(2):302)。综上所述,这六种不同的体外和体内试验有力地支持了bleogen pb1是一种有丝分裂原并发挥egf样生物活性。
[0166]
实施例4:bleogen pb1与egfr相互作用
[0167]
为了表明bleogen pb1的促有丝分裂活性是其与egfr相互作用的结果,使用拉下和中和抗体试验来检查它们的相互作用。生物素-pb1能够拉下egfr,表明它们存在推定的配体-受体相互作用(图4a)。egfr中和抗体(克隆la1)的共同培养阻断了bleogen pb1诱导的hacat细胞增殖(图4b)。这些结果有力地支持了bleogen pb1和egfr之间的相互作用是特异性的,并且增殖作用是依赖于egfr的。
[0168]
实施例5:bleogen pb1的增殖作用与egfr/mek/erk信号通路相关
[0169]
egf激活egfr/mek/erk信号通路,在调节细胞增殖中起重要作用。探讨了bleogen pb1是否会经历类似的egf信号通路以诱导角质形成细胞增殖。我们发现bleogen pb1诱导egfr、mek1/2和erk1/2的磷酸化(图5a、b)。此外,与小分子egfr酪氨酸激酶抑制剂(ag1478)或mek抑制剂(u0126)共同培养显着抑制了hacat细胞中bleogen pb1的增殖作用(图5c、d)。总的来说,这些结果表明bleogen pb1与egfr结合并激活egfr/mek/erk信号通路以触发hacat细胞增殖。
[0170]
已知egf可激活与血清反应元件(sre)结合的转录因子,并启动参与egfr介导的细
胞增殖调节的即刻早期基因的转录,例如c-fos和c-jun(lee et al.(2018)sci.rep.8(1):162)。可以证明bleogen pb1依赖剂量而增加荧光素酶活性,类似于稳定表达的sre-荧光素酶报告基因hacat细胞系中的egf(图5e),表明bleogen pb1刺激sre介导的基因转录。还使用qpcr检测了egf和bleogen pb1诱导的转录反应对egf相关的即刻早期基因表达的影响(brankatschk et al.(2012),sci.signal.5(215):ra21-ra21)。已证实bleogen pb1和egf均显着上调hacat细胞中c-fos和c-jun的mrna表达(图5f)。总之,这一系列实验支持了egf和pb1诱导的信号通路和转录反应的相似性。
[0171]
实施例6:使用ala-和d-氨基酸对bleogen pb1中含有yagqk基序的环4进行位置扫描
[0172]
假设bleogen pb1的环4和典型egfr激动剂的环c中的yxgxk/r基序(x,任何氨基酸;seq id no:3)可能是egfr相互作用的常见“热点”。此前已经证明使用ala或d-氨基酸扫描tgf-αyxgxr基序环c中的y38和r42突变导致egfr亲和力和egf样有丝分裂潜能的显着降低(tam和tam等人,同上)。因此,化学合成了bleogen pb1、yagqk环4的相应“热点”的重点ala和d-氨基酸库。这包括[y25a]pb1、[g27a]pb1、[q28a]pb1和[k29a]pb1的ala置换系列和[y25y]pb1、[a26a]pb1、[q28q]pb1和[k29k]pb1的d-氨基酸置换系列。在两项不同的研究中将每种肽与bleogen pb1进行比较,以使用基于时间分辨荧光能量转移(tr-fret)的竞争性置换测试其对egfr的亲和力,并使用hacat细胞增殖测定测试其有丝分裂潜力。结果表明,bleogen pb1以1720
±
0.075nm的ic50置换生物素-egf,而egf的ic50为31
±
0.016nm(图6a)。通过egfr置换和细胞增殖试验(数据未示出),所有ala类似物均显示egf样生物活性降低。相比之下,两个d类似物[k29k]pb1和[y25y]pb1显示出比bleogen pb1更高的亲和力,ic50分别为27
±
0.050nm和580
±
0.089nm。与bleogen pb1相比,[k29k]pb1的效力大约高出60倍(图6b)。结果表明,生物素-[k29k]pb1和生物素-[y25y]pb1,如之前针对生物素-pb1所述是化学合成的,使用拉下分析与egfr相互作用(数据未示出)。这一数据表明[k29k]pb1和[y25y]pb1比bleogen pb1更具促有丝分裂作用,并且它们的促有丝分裂作用可以被egfr中和抗体(克隆la1)抑制,表明它们的活性是依赖egfr的(图6c)。
[0173]
实施例7:bleogen pb1在使用链脲佐菌素诱导的糖尿病小鼠的切除伤口模型中加速伤口愈合
[0174]
探讨了体外试验中观察到的[k29k]pb1增强的促有丝分裂作用是否也会加速链脲佐菌素(stz)诱导的糖尿病小鼠的伤口愈合。在第1天和第5天通过两次腹膜内注射stz(80mg/kg)在c57小鼠中诱导糖尿病。从第8天开始,定期监测血糖两周。血糖水平高于11.1mmol/l的小鼠被认为患有糖尿病,并接受夹板切除伤口愈合模型(huang et al.(2018),faseb j.33(1):953-964)。结果表明,与非糖尿病小鼠相比,stz诱导的糖尿病小鼠的伤口愈合明显延迟。结果还表明,连续五天用egf(1nmol/伤口)、bleogen pb1(1nmol/伤口)或[k29k]pb1(1nmol/伤口)局部处理的伤口从第4天开始以明显更快的速度愈合,受伤后宏观上与载体对照组相比(图7)。值得注意的是,[k29k]pb1以与egf相似的速率促进伤口愈合,并且比bleogen pb1快得多。这些体内结果与egfr亲和力和促有丝分裂的体外试验结果一致,表明[k29k]pb1与egf具有同等效力。
[0175]
实施例8:bleogen pb1和[k29k]pb1对蛋白水解降解具有超稳定性
[0176]
确定了结构上比egf更紧凑的bleogen pb1和[k29k]pb1是否更具有蛋白水解稳定
性并且对热和酸处理具有抗性,所有三种测试肽bleogen pb1、[k29k]pb1和egf都对酸较稳定,计算值t1/2》500分钟(数据未示出)。然而,观察到bleogen pb1和[k29k]pb1在热处理中明显比egf更稳定,计算值t1/2》500分钟(图8a)。更重要的是,它们还用一组不同的蛋白酶以测试其蛋白水解稳定性。这些包括胃蛋白酶、人血清、胰蛋白酶、链霉蛋白酶和人中性粒细胞弹性蛋白酶(图8b-f)。与egf相比,bleogen pb1和[k29k]pb1在对我们的蛋白酶组的敏感性方面表现出鲜明的对比。bleogen pb1和[k29k]pb1的计算值t1/2在蛋白酶组中》500分钟,在人血清中》800小时。相比之下,egf的t1/2在这组的蛋白酶存在下为0.9到23.5分钟,在人血清中为3.1小时。与bleogen pb1和[k29k]pb1相比,egf在蛋白水解条件下的稳定性至少低100倍。对照肽s-烷基化bleogen pb1的所有二硫键都被碘乙酰胺还原和s-烷基化,在几分钟内被蛋白酶降解,这表明bleogen pb1和[k29k]pb1的结构完整性有助于它们的蛋白水解稳定性。
完整全部详细技术资料下载
当前第1页 1 2
相关技术
- 一种灵芝菌GZ及其应用的制作...
- 一种离子化磺化聚醚醚酮隔膜及...
- 一种耐高温乳液型酸液稠化剂及...
- 一种磷酰基乙酸三乙酯的制备方...
- 一种树脂组合物及其半固化片、...
- 一种高表达CD106和/或C...
- 一种超疏水低析出可降解材料及...
- PIP4K通过非催化依赖性机...
- 一种适合吹塑的耐溶剂ABS复...
- 一种聚(碳酸酯-醚)多元醇分...
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1