免药物筛选快速获得小鼠Rosa26基因定点整合外源基因的方法与流程
本发明属于生物技术基因工程领域,具体涉及一种免药物筛选快速获得小鼠rosa26基因定点整合外源基因的方法。
背景技术:
成簇规律间隔短回文重复(clusteredregulatoryinterspacedshortpalindromicrepeat,crispr)技术是近年来新发展的基因组编辑技术,其通过引导核酸内切酶切割dna靶位点序列,形成dna双链断裂(dnadouble-strandbreak,dsb),从而诱导细胞进行dna损伤修复,然后通过非同源末端连接(non-homologousendjoining,nhej)/同源重组(homology-directedrepair,hdr)途径实现基因组编辑,包括基因敲除(knockout)、基因定点突变及外源基因的定点整合(knockin)。crispr/cas9技术通过人工合成的sgrna序列特异识别靶位点,引导cas9蛋白进行dna双链切割。近些年来,crispr/cas9技术以高效、精准的特点在基因敲除方面得到了广泛应用。
外源基因的定点整合依赖于hdr修复途径,然而细胞内发生dna双链断裂后的主要修复机制是nhej——往往导致单碱基的缺失、插入从而实现基因敲除,这导致了基因knockin的难度远大于knockout,因此在进行基因knockin时选择高效的编辑工具和有效的筛选方法尤为重要。
技术实现要素:
本发明的目的是针对以上要解决的技术问题,提供一种简便、快速、高效、精确的小鼠rosa26基因定点整合外源基因的方法。
为了实现以上目的,本发明提供了一种免药物筛选快速获得小鼠rosa26基因定点整合外源基因的方法,其包括以下步骤:
1)选择小鼠rosa26基因打靶位点;
2)设计靶向rosa26的grna;
3)构建crispr/cas9表达质粒px458-rosa26;
4)构建无启动子的带红色荧光报告基因的同源重组供体载体;
5)细胞转染:将所述crispr/cas9表达质粒px458-rosa26与供体载体共转染进入小鼠es细胞;
6)用流式细胞技术先通过绿色荧光分选出成功转染质粒的细胞进行培养7天后,通过红色荧光进行单细胞分选,经培养获得定点整合外源基因的es单克隆细胞系。
优选地,所述crispr表达质粒的grna的oligo序列如seqidno.2和seqidno.3所示。
优选地,所述步骤3)中,将两条oligo序列混合并稀释至10μm,退火并复性使形成双链结构;退火程序为37℃,30分钟;95℃,5分钟;-5℃/min降至20℃;然后将退火后的双链以1:200比例稀释后,取1μl,加入100ng的px458空载体;再加入bbsi酶使所述px458空载体线性化并加入t4连接酶使带有粘性末端的双链与所述px458空载体连接,得到px458-rosa26质粒。
优选地,所述步骤3)还包括将连接好的所述px458-rosa26质粒转入dh5α感受态中,涂板,扩大培养。
优选地,所述步骤4)的细胞转染条件为:待细胞生长至培养皿90%体积时,弃掉培养基,用pbs洗两次,胰酶消化1min,加入适量血清终止反应;将细胞吹打悬浮,200g离心4min,弃上清,用pbs重悬,轻轻吹打混匀;再次离心并弃上清;每1×106个细胞加入5μgpx458-rosa26质粒并用100μl电极缓冲液重悬;室温孵育5分钟后用1400v、20ms电击2次,然后迅速加入预先放置含20%胎牛血清的dmem的六孔板中培养。
优选地,所述外源基因为外源tyr基因及dsred报告基因。
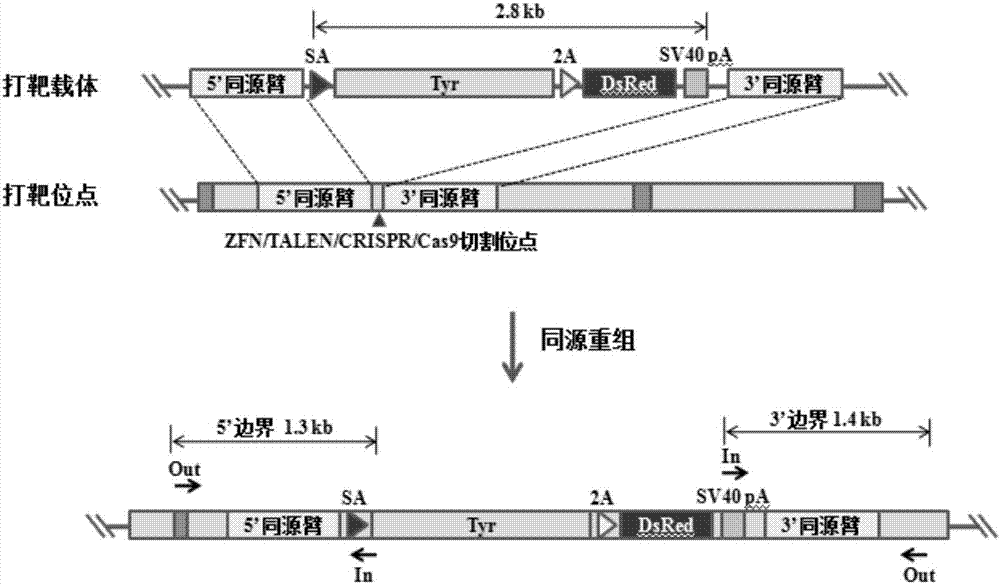
优选地,所述同源重组供体载体为tyr-2a-dsred,其包含位于两端的同源臂,以及位于同源臂中间的外源插入序列,所述外源插入序列包括tyr基因序列与红色荧光报告基因dsred序列,以及剪接受体位点sa。
优选地,所述步骤5)的具体操作为:细胞转染后3d,弃培养基并用pbs洗两次,胰酶消化并200g离心4min,弃上清,pbs洗涤一次,用600μlpbs重悬,经孔径为40μm的细胞网筛筛入流式管中,先后进行绿色荧光和红色荧光的双重荧光筛选。
本发明在rosa26的内含子上运用crispr/cas9基因编辑工具进行了外源tyr基因整合,crispr/cas9的ko活性可达到30%,具有较高的切割效率。对红色荧光的分析结果显示,crispr/cas9的ki效率亦显著较高,达到2.74%,gfp分选后11天细胞ki效率可达到13.6%。
该方法利用绿色荧光和红色荧光的双重荧光筛选,仅仅利用流式细胞技术便可快速鉴定基因编辑ko或ki的效率,更可以快速富集阳性细胞用于进一步实验。相对于传统的正负筛选基因打靶方法,此方法在细胞水平上省去繁琐的筛选过程,更加高效、省时。这种方法不但可以用于前期快速鉴定不同基因工具的定点整合效率,也大大提高了后续获得阳性单克隆es细胞的效率。
经实验发现,crispr/cas9可以更有效地介导外源插入序列定点整合进rosa26内含子中,并且此方法无需经过费时的药物筛选,只需通过流式分选,即可快速高效地获得定点整合的细胞。本发明为今后制作不同的rosa26外源基因定点整合小鼠提供了一种更加精确、简便和高效快速的方法。
附图说明
图1是小鼠rosa26位点同源重组示意图。
图2是小鼠rosa26基因编辑靶位点示意图。
图3是zfn、talen、crispr/cas9基因敲除效率鉴定结果。
图4是zfn、talen、crispr/cas9编辑后的基因测序结果比对。
图5是zfn、talen、crispr/cas9定点整合tyr效率鉴定结果。
图6是利用crispr/cas9构建tyr基因定点整合es细胞的结果,验证pcr扩增中间插入片段。
图7是利用crispr/cas9构建tyr基因定点整合es细胞的结果,验证pcr扩增右侧边界片段。
图8是tyr定点整合的嵌合体小鼠的毛色对比结果。
图9是tyr定点整合的嵌合体小鼠的视网膜对比结果。
具体实施方式
下面结合附图和具体实施例对本发明的技术方案作进一步的说明,但本发明并不限于以下实施例。
在小鼠中,黑色素沉积的类型与数量差异造成了不同的毛色表型。动物的黑色素分为两类:真黑色素(eumelanin)与褐黑色素(pheomelanin),这两种黑色素的合成均依赖于酪氨酸的氧化,而酪氨酸酶(tyr)是催化酪氨酸氧化的关键酶。当外源tyr基因整合进白色小鼠中,成功整合外源tyr的基因的白色小鼠会产生黑色素沉积,产生不同于对照组的毛色表型。因此当构建基因敲入小鼠时,tyr可以作为一个遗传筛选标记,使得可以通过毛色来快速、高效地鉴别knockin小鼠。
在本发明中,选择了公认的safeharbor位点——rosa26的第一个内含子——作为外源基因整合的靶位点。rosa26基因广泛存在于哺乳动物的多种组织中,它转录得到三个不同的非编码rna,对该基因的突变研究表明杂合或纯合的突变均不会引起小鼠的异常表型。因此该位点进行基因敲入可以使转入基因在所有组织和细胞中广泛而稳定的表达,并且对小鼠或细胞的生存无明显影响。
为方便、快捷地获得tyr基因整合的小鼠,构建了tyr-2a-dsred同源重组供体载体(见图1),该载体包含位于两端的同源臂,以及位于同源臂中间的外源插入序列。插入序列包括了要整合的tyr基因序列与红色荧光报告基因(dsred)序列。此外,还有一个剪接受体位点(splicingacceptor,sa),该受点的存在使插入序列被整合到基因组后,在转录后剪接时与5’端外显子连接,保证插入序列可以正常翻译。将该同源重组载体与基因编辑工具zfn、talen、crispr/cas9共转染进入胚胎干细胞(embryonicstemcell,es),目标位点被切割产生双链断链(dsb),通过同源重组介导的修复(hdr)途径,外源序列被正确整合进基因组,dsred基因得以表达,细胞将产生红色荧光。通过流式细胞技术(fluorescenceactivatedcellsorting,facs)可将发出红色荧光的细胞筛选并富集起来,通过进一步培养和鉴定即可得到knockin的es单克隆细胞系。将这些es细胞进一步培养后注射入小鼠囊胚中,再通过胚胎移植技术将这些囊胚植入假孕母鼠的子宫,即可获得部分细胞含有外源tyr基因插入的嵌合体小鼠。
1材料和方法
材料
c2c12细胞(小鼠成肌细胞)(可商购,由本实验室保存),昆明小鼠(kunmingmice,km)来自中山大学实验动物中心。zfn质粒和talen由美国sigma公司合成,px458质粒购自addgene公司。引物及oligo序列由上海生工生物技术公司合成。
方法
rosa26基因靶位点选择
选取rosa26基因第1200位碱基附近序列作为基因编辑的靶位点。在该位点附近可以同时设计1对zfn、2对talen以及1条crisprgrna(seqidno.1,5’-agtctttctagaagatgggc-3’),三者的切割位点相近(见图2)。
zfn与rosa26内含子的结合区域如图2所示,两段zfn结合序列中间的序列为切割位点。zfn表达质粒由上海生工生物技术公司合成。
talen与rosa26内含子的结合区域如图2所示,分别设计了两对talen表达质粒,为talena和talenb。质粒由上海生工生物技术公司合成。
crispr/cas9基因编辑质粒oligo的设计
选取pam位点前20个碱基作为grna结合位点。设计oligo时在5’端加上bbsi内切酶的粘性末端。
crispr/cas9表达质粒的grnaoligo序列如下所示:
正义链(top):5’-caccgagtctttctagaagatgggc-3’;(seqidno.2)
反义链(bottom):5’-aaacgcccatcttctagaaagactc-3’。(seqidno.3)
oligo由广州艾基生物技术有限公司合成。
px458载体的构建
将两条oligo序列混合并稀释至10μm,退火并复性使形成双链结构。退火程序为37℃,30分钟;95℃,5分钟;-5℃/min降至20℃。将退火后的双链以1:200比例稀释后,取1μl,加入100ng的px458空载体。加入bbsi酶使上述px458空载体线性化并加入t4连接酶使带有粘性末端的双链与px458空载体连接,得到px458-rosa26质粒。将连接好的px458-rosa26质粒转入dh5α感受态中,涂板,挑选单克隆测序,比对成功后扩大培养,抽提去内毒素质粒备用。
细胞转染
选用15代以内的细胞,待生长至90%汇合度时,弃掉培养基,用pbs洗涤两次,胰酶消化1min,加入适量血清终止反应。将细胞吹打悬浮,转移到ep管里,200g离心4min,弃上清,用pbs重悬,轻轻吹打混匀,取10μl用细胞计数仪计数。再次离心并弃上清。每1×106个细胞加入5μg质粒并用100μl电极缓冲液重悬。室温孵育5分钟后用1400v、20ms电击2次,然后迅速加入预先放置含20%(v/v)胎牛血清的dmem的六孔板中。培养24h后换液。
t7e1实验
设计t7e1引物,使pcr扩增打靶位点两侧共约500bp的序列(seqidno.4),并使打靶位点位于pcr产物序列约100-200bp的位置。
rosa26编辑位点扩增引物如下所示:
rosa26-f:gcacgtttccgacttgagtt;(seqidno.5)
rosa26-r:cctcccattttccttatttgc。(seqidno.6)
t7e1内切酶可以识别dna双链上的环状突起并进行双链切割。由于dna双链在编辑工具的切割后进行nhej(非同源性末端接合)修复后,往往会产生插入或缺失(indel),因此经变性-退火后的产物由于来自不同基因编辑的dna链与野生型的dna链之间会产生错配,形成环状突起,被t7e1内切酶识别切割。因此,依据t7e1酶的切割效率,可以估算出基因编辑产生indel的效率。
收集细胞并提取基因组,pcr扩增,切胶回收。95℃变性10min,-5℃/min缓慢退火至25℃。取200ng变性-退火产物,在20μl体系中加入0.5μlt7e1内切酶。37℃酶切30min,10%(v/v)page凝胶120v、90min电泳,eb染色5min后紫外观察条带,根据条带的强弱比较切割效率的高低。
流式分选
细胞转染后3d,弃培养基并用pbs洗两次,胰酶消化并200g离心4min,弃上清,pbs洗涤一次,用600μlpbs重悬,经孔径为40μm的细胞网筛筛入流式管中,上机分析荧光比例与强度并用另一支流式管收集带有荧光的细胞。对于要分选培养单克隆的细胞,流式细胞仪设置为96孔板,每孔1个细胞,用含有20%血清的完全培养基的96孔板收集,培养7d后换液并进一步扩大培养。
嵌合体小鼠制备
促排四周龄母小鼠,并与公鼠合笼,检查交配栓。3.5d后引颈法处死母鼠并收集输卵管和子宫中的胚胎。转移到覆有石蜡油的m16培养基内,37℃培养。
将分选并扩培后的es单克隆细胞用胰酶消化成单细胞悬液,并转移到m2培养基液滴内,同时将培养好的囊胚转移到液滴内。用显微注射系统将es细胞注射到囊胚腔内,每个囊胚注射10-12个es细胞。37℃培养1-3h,待囊胚腔恢复后,移入代孕母鼠子宫中培养。
2结果与分析
2.1zfn、talen、crispr/cas9基因组切割效率鉴定
将构建好的zfn、talen和crispr/cas9表达质粒分别电转染c2c12细胞,24小时后换液,待转染细胞长满六孔板,消化细胞并提取基因组。用rosa26-t7e1引物扩增,切胶回收目标条带并进行t7e1酶切实验,以检测三种基因编辑工具在rosa26目标位点的切割效率。pcr扩增的条带大小约为500bp,与预期一致;酶切实验显示未转染的c2c12细胞条带未被切割,转染zfn的细胞的pcr条带也未被切割。转染talen-a的细胞pcr条带也未发生切割而转染talen-b的则切割成功。转染crispr/cas9的细胞的pcr条带也被切开,并且割后产生的两条条带比talen的更亮,因此初步判断crispr/cas9在该位点的基因编辑效率要优于talen和zfn(见图3)。将pcr扩增产物构建ta克隆,并各挑选50个单克隆进行测序,计算发生碱基缺失或插入或突变的比率,得到zfn的编辑效率为4.1%,talen-a的编辑效率为4.2%,talen-b的编辑效率为24%,crispr的编辑效率为30%(见图4)。因此,crispr/cas9在该位点具有相对较高的编辑效率。
2.2zfn、talen、crispr/cas9整合外源tyr基因效率鉴定
将zfn、talen和crispr/cas9质粒分别与同源重组供体载体(donor)共转入c2c12细胞,24h后换液,在荧光显微镜下可观察到带有红色荧光的细胞,表明重组片段可能已正确整合到目标位点。待转染细胞长满六孔板后,用流式细胞仪进行荧光分析。crispr/cas9编辑的细胞dsred阳性率可达到2.74%,显著高于zfn和talen编辑的细胞。接下来利用crispr的载体质粒px458上带有gfp荧光蛋白的特点对这些细胞进行进一步的富集。将共转染crispr和donor后培养24小时的细胞进行流式分选,筛选出gfp阳性的细胞,将这些细胞继续培养,待长满后再次用流式细胞仪分选出dsred阳性的细胞。提取基因组并分别对两侧边界序列及中间的插入片断进行巢式pcr扩增,琼脂糖凝胶电泳检测片段大小并回收测序。pcr成功扩增出了3’端跨越插入序列和rosa26内含子的边界片段,大小约1.4kb,符合预期(见图5)。测序结果进一步表明插入片段已正确整合到位于rosa26内含子上的目标位点。
2.3crispr/cas9构建tyr基因定点整合es细胞
由于crispr/cas9具有较强的基因定点整合效率,因此选用crispr/cas9作为基因编辑工具来获得外源tyr插入的es细胞。将构建好的px458-rosa26质粒与donor共转入es细胞,24h后换液,荧光显微镜下可见有部分细胞同时表达gfp和dsred。待细胞长满后流式细胞仪筛选带有绿色荧光的细胞。继续培养至长满六孔板,用流式细胞仪分选表达dsred的单个细胞至96孔板的每一孔中,3天后显微镜下可见部分细胞分裂形成单克隆细胞群。继续培养这些单克隆并传代使之可铺满六孔板,取一部分细胞提取基因组并进行pcr扩增。在得到的5个单克隆中,有3个克隆成功扩增出了中间的tyr基因插入序列(见图6),其中2个克隆成功扩增出了右侧边界片段(见图7)。切胶回收并测序,结果符合预期,说明成功得到了tyr基因敲入的es细胞。
2.4构建外源tyr基因整合嵌合体小鼠
成功整合外源tyr基因的es细胞经培养后,注射入囊胚并将囊胚移植到代孕母鼠输卵管内,同时移植注入未转染的es细胞的囊胚作为对照。实验组的母鼠成功产出嵌合体小鼠一只,该小鼠的毛色呈现部分灰色,而同窝非嵌合体鼠和对照组嵌合体小鼠均呈白色(见图8)图中右2为tyr嵌合体实验小鼠。该小鼠的视网膜较对照组亦呈现更深的黑色(见图9),图中左侧为嵌合体对照小鼠,右侧为tyr嵌合体实验小鼠。
本发明在rosa26的内含子上,分别运用zfn、talen、crispr/cas9基因编辑工具在同一位点进行了外源tyr基因整合。在该位点上,三种基因编辑工具均表现出了靶向切割活性,其中zfn的dna双链切割活性为4.1%,其中一对talen可达到24%,而crispr/cas9可达到30%,具有更高的切割效率。对红色荧光的分析结果显示,crispr/cas9的ki效率亦显著高于zfn和talen,在c2c12细胞中,crispr/cas9的ki效率达到2.74%,为zfn的9.7倍,talen的8.3倍。gfp分选后11天细胞ki效率可达到13.6%,具有明显的优势。
该方法利用绿色荧光和红色荧光的双重荧光筛选,仅仅利用(流式细胞技术)facs技术便可快速鉴定基因编辑ko或ki的效率,更可以快速富集阳性细胞用于进一步实验。相对于传统的正负筛选基因打靶方法,此方法在细胞水平上省去繁琐的筛选过程,更加高效、省时。这种方法不但可以用于前期快速鉴定不同基因工具的定点整合效率,也大大提高了后续获得阳性单克隆es细胞的效率。
对ki细胞基因组的巢式pcr扩增得到了3’端边界序列和中间插入序列的条带,证明了外源基因的成功整合。左侧插入序列的扩增无法得到条带,原因可能是左侧同源臂有大量gc重复序列,这同样增加了扩增的难度。
最后,成功得到了整合了tyr基因的嵌合体小鼠,该小鼠相对于非嵌合体小鼠具有明显的毛色区别,并且眼睛也呈现更深的黑色。通过毛色可以更加直观、快速地鉴别目标外源基因整合小鼠,这为今后制作rosa26定点整合小鼠提供了一种简便高效的方法。
序列表
<110>中山大学
<120>免药物筛选快速获得小鼠rosa26基因定点整合外源基因的方法
<160>6
<210>1
<211>20
<212>dna
<213>人工序列
<220>
<221>misc_feature
<400>1
agtctttctagaagatgggc20
<210>2
<211>25
<212>dna
<213>人工序列
<220>
<221>misc_feature
<400>2
caccgagtctttctagaagatgggc25
<210>3
<211>25
<212>dna
<213>人工序列
<220>
<221>misc_feature
<400>3
aaacgcccatcttctagaaagactc25
<210>4
<211>200
<212>dna
<213>人工序列
<220>
<221>misc_feature
<400>4
gcacgtttccgacttgagttgcctcaagaggggcgtgctgagccagacctccatcgcgca60
ctccggggagtggagggaaggagcgagggctcagttgggctgttttggaggcaggaagca120
cttgctctcccaaagtcgctctgagttgttatcagtaagggagctgcagtggagtaggcg180
gggagaaggccgcacccttctccggaggggggaggggagtgttgcaatacctttctggga240
gttctctgctgcctcctggcttctgaggaccgccctgggcctgggagaatcccttccccc300
tcttccctcgtgatctgcaactccagtctttctagaagatgggcgggagtcttctgggca360
ggcttaaaggctaacctggtgtgtgggcgttgtcctgcaggggaattgaacaggtgtaaa420
attggagggacaagacttcccacagattttcggttttgtcgggaagttttttaatagggg480
caaataaggaaaatgggagg500
<210>5
<211>20
<212>dna
<213>人工序列
<220>
<221>misc_feature
<400>5
gcacgtttccgacttgagtt20
<210>6
<211>21
<212>dna
<213>人工序列
<220>
<221>misc_feature
<400>6
cctcccattttccttatttgc21
- 一种关于李斯特菌的电转化方法...
- 处理牛体细胞克隆胚的方法与流...
- 一种人正常CHCHD2过表达...
- SOD2慢病毒载体、构建方法...
- 一种携带iCasp9自杀基因...
- 一种共抑制油菜FAD2和FA...
- 对农杆菌介导转化法获得的红掌...
- AtGA2ox1基因在调节植...
- 三七几丁质酶基因PnCHI1...
- 一种适用于植物的大规模基因组...
- 还没有人留言评论。精彩留言会获得点赞!
- 一种C5aR基因敲除小鼠cGVHD模型的建立方法与制造工艺
- 基因工程鼠保种隔离器的制造方法
- 一种幽门螺杆菌基因无痕敲除的方法
- 一种多重pcr快速检测稻曲菌交配型基因的方法
- 一种PS1基因条件性敲除flox大鼠的制备方法
- 一种乙二胺四乙酸二酸酐修饰草酸脱羧酶的方法
- 杀香鱼假单胞菌ExoU基因敲除突变株及其应用
- 一种ApoE试剂盒、引物及其用图
- 基因vii型新城疫病毒株、其疫苗组合物、制备方法及应用
- 一种快速获得基因敲除细胞株的方法
- 一种定点敲除几丁质合成酶基因的方法
- 一种Fbxo40基因敲除猪的制备方法
- GABAergic神经元条件性敲除基因PGC-1α小鼠模型及其构建方法
- 基因工程鼠保种隔离器的制造方法
- GABAergic神经元条件性敲除基因PGC-1α小鼠模型及其构建方法
- 基因工程鼠保种隔离器的制造方法
- 水稻OsCSA基因的应用及其定点敲除方法
- 一种建立cyp2c11基因敲除大鼠模型的方法
- 一种建立cyp2d1基因敲除大鼠模型的方法
- 通过CRISPR/Cas9技术得到敲除铁调素基因斑马鱼的制备方法
- 一种利用混合水生植物制备姬菇栽培基料的方法与制造工艺
- 一种大菱鲆增食欲素Orexin基因、重组蛋白及其制备方法与制造工艺
- 一种小型啮齿类实验动物航空运输装置的制造方法
- 一种抗草甘膦转基因籼稻的制备方法与制造工艺
- 一种桐油基多元醇及其制备方法
- 基因工程鼠保种隔离器的制造方法
- 一种水基防锈剂的制备方法
- eGFP标记GPR120阳性细胞的小鼠模型构建方法
- 一种新型转基因复合物及其制备方法与应用
- 水基油脂清洗剂及其制备方法
- 鸭肠炎病毒gE、gI双基因缺失病毒株的构建及其应用
- 一种5基因缺失伪狂犬病重组病毒活疫苗及其制备方法
- 一种猪链球菌2型五基因缺失株及应用
- 一种小鼠色素缺失动物模型的制备方法
- 一种检测vhl基因大片段缺失的杂交探针及检测方法与试剂盒的制作方法
- 鼠spata16基因多克隆抗体及其制备方法
- 鼠Bpifb5基因多克隆抗体及其制备方法
- 鉴别PRV强毒和PRV gG、PRV gE基因缺失疫苗毒的PCR引物的制作方法
- 猪链球菌SBP_bac_5基因缺失株及其构建方法与应用
- 一种缺失sfp基因的生防菌及其发酵工艺和应用
- 一种针对大小鼠的针灸器械的制作方法
- 一种大小鼠代谢笼的制作方法
- 一种大小鼠的代谢分离器的制作方法
- 多功能大小鼠实验台的制作方法
- HpychH4内切酶鉴定TERF1基因rs3863242多态性的制作方法
- 一种变换核酸基因型的方法
- 一种慢羽公鸡基因型的检测方法
- 一种用于检测牛gart突变基因型的引物对和检测方法
- 试剂盒及确定待测dna样本预定snp位点的基因型的方法
- 基因型解析装置以及基因型解析方法
- 一种鉴定水稻垩白主效基因的基因型的分子标记及其应用
- 基于ssr基因型生成果树ssr分子身份证号码的方法
- 高粱抗白粉病基因快速鉴定的制作方法
- 谷子抗白粉病基因快速鉴定的制作方法